灵芝群体交配基因型分析发表时间:2023-08-24 11:32
Chinese Agricultural Science Bulletin 灵芝群体交配基因型分析 陈裕新1,2,夏志兰1,2,刘 鹏1,2,康信聪1,2,洪亚辉3,刘东波1,2 (1 湖南农业大学园艺园林学院,长沙 410128;2 国家中医药管理局亚健康干预技术实验室,长沙 410128; 3 湖南农业大学生物技术学院,长沙 410128) 摘 要:研究灵芝群体交配基因型,并与酯酶同工酶试验相比较,探讨它们在群体亲缘关系分析上的异同点。采用OWE-SOJ 技术对24 个灵芝菌株的单孢分离菌株进行标准交配型鉴定,及群体间交配基因型的测定,并结合酯酶同工酶试验对群体间的亲缘关系进行分析。鉴定出7 大类交配基因型,并发现A 因子含有4 个等位基因,B 因子含有4 个等位基因,及1 个特殊的A 混合基因,4 类交配型出现了一定程度的偏分离。酯酶同工酶试验检测出了28 条不同酯酶酶谱带,在相似系数为0.73214 时,样品被分为9 大类。比较交配基因型分析与酯酶同工酶试验结果,发现两者具有很高的相似性。交配基因型测定可以作为分析菌株间差异和鉴定品种的一种重要补充手段。 关键词:酯酶同工酶;OWE-SOJ 技术;交配型;菌落形态 中图分类号:Q36文献标志码:A论文编号:2011-3236 Mating Genotype Analysis of Ganoderma lucidum Populations Chen Yuxin1,2, Xia Zhilan1,2, Liu Peng1,2, Kang Xincong1,2, Hong Yahui3, Liu Dongbo1,2 (1Horticulture and Landscape College, Hunan Agricultural University, Changsha 410128; 2State Administration of Traditional Chinese Medicine, State Key Laboratory of Subhealth Intervention Technology, Changsha 410128; 3College of Bioscience and Biotechnology, Hunan Agricultural University, Changsha 410128) Abstract: The mating genotype was studied and compared with esterase isozyme of Ganoderma lucidum populations between groups in order to clarify their differences in genetic relationship analysis. OWE-SOJ technique was applied to identify standard mating types and determinate mating genotype between groups of monokaryons isolates from 24 G. lucidum stains. Genetic relationships were analyzed by combined group mating genotype determination with esterase isozyme assay. The results showed that all strains of G. lucidum could be divided into 7 large groups of the mating genotype. 4 alleles of A factor, 4 alleles of B factor and one mixed alleles of A factor were found in this study. Distorted segregation ratio among monokaryon mycelia of G. lucidum had been observed in 4 kinds of mating types to some extent. 28 different types of enzyme bands were determined in esterase isoenzyme test. 28 strains of G. lucidum could be divided into 9 large groups through the cluster analysis when the genetic similarity coefficient was 0.73214. Comparing mating genotype analysis with esterase isozyme analysis, it showed great similarity. Mating genotype analysis could be an important supplementary method to strains identification and genetic diversity research. Key words: esterase isozyme; OWE-SOJ technique; mating type; colony morphology 基金项目:国家科技支撑计划“中药产业区域发展及特色产品研究开发”(2006BIA06A20);科技部科技人员服务企业行动计划“珍稀食用菌系列产品开发”(2009GJD20012)。 第一作者简介:陈裕新,男,1987 年出生,广西来宾人,在读硕士,研究方向为药用植物资源工程。通信地址:410128 湖南长沙市湖南农业大学生命科学楼亚健康干预技术实验室,Tel:0731-84635293,E-mail:hotdogchenyuxin@163.com。 通讯作者:刘东波,男,1970 年出生,湖南长沙人,副教授,硕士生导师,博士,研究方向为中药资源与开发。通信地址:410128 湖南长沙市湖南农业大学生命科学楼亚健康干预技术实验室,Tel:0731-84635293,E-mail:chinasaga@163.com。 收稿日期:2011-11-07,修回日期:2012-02-17。 · 214 · 引言 中国农学通报http://www.casb.org.cn 么可以构成23328 种不同A、B 因子的组合[4]。除此之 灵芝(Ganoderma lucidum)属真菌界,担子菌亚门, 层菌纲,非褶菌目,灵芝科,灵芝属,为大型的药用真菌。有着“仙草”的美誉,是名贵的药材,被广泛应用于药品和食品中。灵芝是一种典型的四极性异宗结合的高等担子菌,属于双因子不亲和性系统,由不亲和性因子A 和B 组成[1]。 对于四极性异宗结合菌而言,同一品种的各菌株理论可亲和率为25%。但是,高等担子菌均存在广泛的复等位基因现象,即一个决定交配型的基因位点上出现多个等位基因。例如,组成A、B 因子的亚基α、β 具有一系列的复等位基因,当α或β 2 个亚基的等位基因发生变化时,会形成新的A 因子或B 因子[2]。复等位基因的存在,提高了自然群体中异系间的单孢随机配对 亲 和 率 [3]。 目 前 发 现 ,裂 褶 菌 (Schizophyllum commune)的Aα和Aβ等位基因分别为9 和32 个,Bα和 Bβ的等位基因都为9 个,则理论上A 因子的复等位基因数位达到288 个,B 因子的等位基因数位有81 个,那 外 ,还 发 现 平 菇 (Pleurotus Ostreatus) [5]、金 针 菇(Flammulina velutipes)[6-7]、香菇(Lentinula edodes) [8] 等的A、B 因子也有多个复等位基因。 复等位基因的存在,导致了灵芝种质资源的多样性,也是其杂交育种的基础,因而,准确鉴定出不同品种的灵芝交配基因型,将为灵芝和其他食药用真菌的遗传研究、选择亲本提供可靠的理论依据。目前,对于灵芝栽培和药理功效等方面的研究甚多,但对其基础遗传学方面的研究,尤其是交配基因型研究并未见报道。笔者以24 个灵芝菌株为样本,研究了灵芝交配型等位基因的数目,并与酯酶同工酶进行对比,探讨交配型测定在分析菌株间差异和品种鉴定上的应用,为灵芝种质遗传多样性的研究提供参考。 材料与方法 实验材料 供试菌株 24 个灵芝菌株的来源见表1,均由湖南农业大学国家中医药管理局亚健康干预技术实验室 表1 供试菌株来源
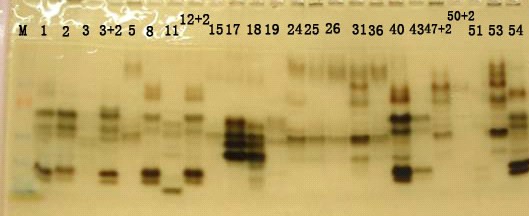
陈裕新等:灵芝群体交配基因型分析 · 215 · 保存。 培养基 PDA 加富培养基:马铃薯(去皮)20%,葡萄糖2%,KH2PO4 0.3%,MgSO4 0.15%,酵母膏0.2%,蛋白胨0.2%,琼脂2%,pH 6.5~7.0。 橡树木屑浸汁琼脂(OWE):5% 橡树木屑浸汁 200 mL,琼脂20 g,加蒸馏水至1000 mL。 榨桔汁琼脂(SOJ):新鲜榨桔汁200 mL,琼脂20 g, 加蒸馏水至1000 mL。 实验方法 酯酶同工酶分析具体参照文献[9]。单孢分离采用常规稀释分离法——平板涂布法,具体参照文献[8]。常规交配型测定采用3 轮杂交系统试验方法,具体参照文献[10]。标准交配型测定经过常规交配型测定后,再采用OWE-SOJ 技术进行标准交配型测定,具体参照文献[11]。菌株间群体交配型基因的测定:随机 选取1 个菌株,将其4 个交配型单核菌株设为基准测试菌株,与其他菌株的4 个交配型单核菌株进行两两交配反应,镜检锁状联合的有无,再根据交配反应的结果进行分析,推测待测菌株交配基因型;如本次没能推测出交配基因型的菌株,再进行下一轮的交配反应,直至所有菌株交配基因型推测完毕。 结果与分析 酯酶同工酶结果分析 电泳结果如图1 所示,24 个菌株共检测出28 条迁移率(Rf)不同酶谱带,Rf 分布在 0.0508~0.9153 之间。 0.5631、0.6911、0.7552、0.5111 4 组条带为大部分菌株共有酶谱带;样品的相似系数范围为0.536~0.964,最大相似系数出现在12+2 号和40 号,25 号和36 号之间,它们之间可能出现同物异名现象;最小相似系数出现在11 号和19 号,11 号和50+2 之间。43 与54 号原名都
图1 24 个灵芝菌株的酯酶同工酶电泳图谱 为北韩灵芝,但在同工酶分析中却只有0.679 的相似系数,应是同名异种的现象。 用 UPGMA 聚类法对 24 株灵芝样品菌株进行聚类分析,获得聚类树形图如图2,可以看出大约在相似系数为 0.73214 时,样品被分为 9 类;1、5、15、26、25、 26、31、17、18 共 9 个菌株聚为一大类( 记作 I 类); 47+2、50+2 共2 个菌株聚为一类(记作II 类);2、12+2、 40、3+2、8、43、51 共 7 个菌株聚为另一大类(记作 III 类);其余6 个菌株各自独成一类。25 与36 号菌株以 及12+2 与40 号菌株的名称各不相同,但聚类分析表 明,它们的遗传相似系数很高,可能为同一菌株;而43 与54 号原名都为北韩灵芝,但在聚类分析中,它们只有较小的遗传相似系数,应是同名异种的现象。 标准交配型结果分析 标准交配型结果分析如表2 所示,在标准交配型鉴定中,使用 OWE-SOJ 技术检测到无锁状联合的绒毛状菌落(A=B≠,图3),但此类交配反应中的部分组 合能镜检到假锁状联合。假锁状联合中形成的钩状分枝与次末端细胞不能融合,导致产生假锁状联合菌丝 (图4)。而OWE-SOJ 技术检测到无锁状联合的栅栏状菌落中,也没有发现假锁状联合现象(A≠B=,图5)。 24 个菌株共分离单核菌株437 个,从供试菌株内 各交配型分布看,a(AⅠBⅠ)与b(AⅡBⅡ)之和数量上明显多于c(AⅠBⅡ)与d(AⅡBⅠ)之和,其总数和比为a(AⅠBⅠ): b(A Ⅱ B Ⅱ):c(A Ⅰ B Ⅱ):d(A Ⅱ B Ⅰ) = 131:138:75:93=1:1.05: 0.57:0.71,4 类交配型分离中在一定程度上偏离1:1:1:1 的分离比。 2.3 群体交配基因型的测定结果分析 先后经过6 轮的交配测定,结果如表3,24 个供试菌株的交配基因型划分为7 大类,共测定出A 因子含有4 个等位基因,B 因子含有4 个等位基因,及1 个特殊的A 混合基因。A1A2B1B2 和A1A3B1B2 交配基因型在供试菌株中占有很大优势,分别都达到33.3%,其余5 类交配基因型占有小量比例。从遗传进化看,占 · 216 · 类 类 类 中国农学通报http://www.casb.org.cn 1 5 15 26 25 36 31 17 18 47+2 50+2 19 24 2 12+2 40 3+2 8 43 51 3 53 54 11 0.660.71 0.810.95 Coefficient 图2 24 个灵芝的酯酶同工酶遗传聚类图
图3 无锁状联合的绒毛状菌落(A=B≠)
图4 假锁状联合(箭头所指)(400×) 有优势比例的交配基因型是进化的主流,占劣势比例的交配基因型多为变异菌株或进化的分支,遗传关系较远。 图5 无锁状联合的栅栏型菌落(A≠B=) 结论与讨论 本研究对24 个灵芝菌株的酯酶同工酶进行分析, 结果显示大约在相似系数为0.73214 时,样品被分为9 类。而群体交配型基因测定显示24 个灵芝菌株划分 为7 大类。比较群体交配型基因与酯酶同工酶分析, 发现占有很大优势的交配型A1A2B1B2 主要分布于I 类中(G.l0050 + 2 菌株除外),而另一大类交配型A1A3B1B2 主要分布于 III 类中(G.l0047 + 2 菌株除外),G.l0019、G.l0054 号菌株和具有A 特殊混合交配型基因的G.l0011 号菌株分别分属于各自聚类中。由此可以得出,交配型分析与酯酶同工酶分析结果具有很高程度上的相似性和一致性,因此,交配基因型测定可以作为分析菌株间差异和鉴定品种的一种重要补充手段。 陈裕新等:灵芝群体交配基因型分析 表2 24 个灵芝菌株标准交配型测定结果 · 217 ·
表3 24 个灵芝菌株群体交配基因型测定结果 菌株编号 G.l0001、G.l0005、G.l0015、G.l0025、G.l0026、G.l0031、G.l0036 、G.l0050+2 G.l0002、G.l003+2、G.l0008、G.l0012+2、G.l0040、G.l0043、G.l0047+2、G.l0051 G.l0003、G.l0024、G.l0053 G.l0017、G.l0018 G.l0019 G.l0054 G.l0011 交配基因型 A1A2B1B2 A1A3B1B2 A2A3B1B3 A1A3B1B4 A1A3B1B3 A1A4B3B4 (A1A2)A3B2B3 在食用菌的有性生殖过程中,交配型基因对单核体菌丝性别控制和双核体菌丝的形成起着决定性作用。灵芝为四极性交配系统,即由2 个独立分配的因子(通常称为A、B 因子)控制交配。其中,A 因子通过 HD1-HD2 异源二聚体调控细胞质融合、细胞核配对、核分裂和锁状细胞形成等发育过程;B 因子则编码信息素及其受体,控制核迁移[12]。在生成异核体时,只有 A、B 因子都处于异等位基因状态时,才能形成稳定异 · 218 · 中国农学通报http://www.casb.org.cn 核体,完成有性生活史。若只有B 位点处于异等位基因状态,A 位点处于等位状态(即A=B≠),核迁移不能稳定下来,钩状分枝与次末端细胞不能融合,导致产生假锁状联合菌丝[11,13],这与本研究的试验结果得出的观点一致。但是,在A=B≠组合中,只有部分产生假锁状联合,并非全部,这与林芳灿[11]等在香菇交配型研究中结果相似,但其具体缘由还值得进一步探讨。 本试验中,灵芝菌株还出现了担孢子交配型因子偏分离现象,这在其他真菌中也有同样发现。如香菇[14]、金针菇[15]等都有报道。对于这种现象,Kawa bata等[16]认为交配型比例偏离与细胞质对核的选择能力有关系,也与A、B 交配型因子连锁的具致死效应的某些生育抑制因子有关。而Raper[17]则指出,出现这种偏分离是与B 因子有关,B 交配型因子影响来自异核体细胞分离的细胞核的存活力,这是不对称核比例现象出现的遗传基础。Kay 等[18]认为,平菇双核菌丝体细胞经原生质体单核化后,2 种核比例的偏离可能与一些形式的核选择有关。灵芝出现这种现象的原因是否与其他真菌一样,还值得深究。 OWE-SOJ 技术是美国学者[19] 在 20 世纪 90 年代初开发并首先应用在密环菌(Armillaria mellea)交配反应鉴定上的一门新技术。其技术优点是实验结果直观,操作简便,缺点为实验周期较长,但是单独运用不能将相同A 的反应区别开来,因此必须结合常规交配型分析,才能将4 类交配型准确区分开来。 参考文献 林志彬,赵大明,许瑞祥,等.灵芝的现代研究(第三版)[M].北京大学医学出版社,2007:9. Cao H. Genetic analysis on the incompatibility factor characteristics in a bipolar basidiomycetes Pholiota nameko[D]. Tottori,Japan: Tottori University,1998. Miles P G. Chang S T. The collection and conservation of genes of Lentinus. Proc. Intil. S-ym. Scientific and technical Aspects of Cultivating Edible Fungi[M].Amsterdam: Elsevier Science Publisher B.V,1987:227-232. Koltin Y. Genetics and Morphogenesis in the Basidiomycetes[M]. New York: Academic Press, 1978:31. 程莉.平菇交配型因子组合的准确鉴定[M].武汉:华中农业大学, 2007. Takemaru T, Suzuki M, Migski N. Isolation and genetic of auxotrophic mutants in Flmmulina velutipes [J].Trans . Mycol. Soc. Japan,1995,36:152-157. Casselton L A, Kues U. The mating tpye genes of Homobasidiomycetes[A].K Esser,P A Lemke. The Mycota I Growth Differentiation and Sexuality[C]. Berlin, G. Berlin, Gmany: Spinger Verlag,1994:307-321. 林芳灿,张树庭.中国香菇栽培菌株不亲和性因子的分析[J].华中农业大学学报,1995,14(5):459-466. 秦俊哲,谷坤睿,陈合.14 个灵芝菌株的酯酶同工酶分析[J].微生物学通报,2005,32(4):20-24. 董洪新,蔡德华,李玉.猪肚菇担孢子交配型的分析[J].微生物学通报,2010,37(11):1617-1620. 林芳灿,汪中文,熊再明,等. OWE-SOJ 技术及核迁移试验在香菇交配型因子鉴定中的应用[J]. 华中农业大学学报,2000,19(6): 573-576. 李莉云,刘国振,刘丽娟.担子菌交配型基因的克隆及功能研究进展[J].生物工程进展,1999,19(1):19-23 彦培璐,姚方杰,孙露,等.金顶侧耳不亲和性因子多样性的研究[J]. 中国食用菌,2011,30(1): 38-41. Cueng Shuiming, Lin Fangcan, Xu Xuefeng, et al. Genetic analysis of segregation distortion of mating-type Factors in Lentinula edodes [J]. Progress in Natural Seience,2005,15(8),84-88. 江玉姬,王秀全,刘维侠,等. 金针菇不亲和性因子研究[J].食用菌学 报,2000,16(4):16-19. Kawa bata H, Magae Y, Sasaki T, et al. Mating type analysis of monokaryons regenerated from protoplasts of Flammulina velutipes [J]. Trans Mycol Soc Japan, 1992,33(2):243-247. Raper C A. B mating type genes influence survival of nuclei separated from heterokaryons of Schizophllum[J]. Exs Mycol,1985, 9:149-160. Kay E, Vilgalys R. Spatial distribution and genetic relationships among individusls in a natural population of the oyster mushroom Pleurotus ostreatus[J]. Myeologia,1992,84:173-182. Darmono T W, Burdsall H H. Morphological characteristics of incompatibility reactions and evidence for nuclear migration in Armillaria mellea[J].Mycologia, 1992,84:367-375. |